Durante la primavera de 2011 se ha llevado a cabo en Álava un estudio destinado a actualizar el conocimiento del área de distribución y estimar el tamaño de la población reproductora de picamaderos negro (Dryocopus martius). Aunque la información publicada sobre la especie en los últimos atlas de aves nidificantes e invernantes (Simal & Herrero, 2003; Gainzarain, 2006; figura 1) sugería ya una expansión del área de distribución “tradicional” de Pirineos y Cordillera Cantábrica, en los últimos años ha tenido lugar un incremento considerable del número de citas en zonas intermedias (Burgos y País Vasco, fundamentalmente). Documentar este proceso de expansión ha sido el objetivo del trabajo.
La metodología utilizada consistió en la realización de itinerarios por parte de observadores cualificados, atravesando zonas forestales (hayedos y pinares de pino silvestre y laricio) con el fin de 1) obtener contactos visuales o auditivos con ejemplares, y 2) registrar indicios o rastros de alimentación atribuibles a la especie. El trabajo de campo se efectuó principalmente durante el mes de marzo y la primera quincena de abril, periodo durante el cual se supuso que la detectabilidad de la especie sería mayor. No se emplearon reclamos.
El área de estudio ha sido Álava junto con los enclaves de Treviño (Burgos) y Orduña (Bizkaia). Con ayuda del Mapa de hábitats EUNIS del País Vasco (Gobierno Vasco, 2005), en cada cuadrícula UTM de 100 km2 se identificaron las manchas existentes de las formaciones arboladas mencionadas anteriormente. Las cuadrículas fueron adjudicadas a los participantes, estandarizando el esfuerzo de campo a realizar en cada una de forma aproximadamente proporcional a la superficie ocupada por las manchas potenciales. Así, se planteó la realización de un mínimo de 5 medias jornadas (4-5 horas de duración) en aquellas cuadrículas con mayor superficie boscosa (29 a 37 km2), 2 en las de menor (2 a 6 km2), y 3-4 en las que presentaban coberturas intermedias. Del total de 50 cuadrículas en el área de estudio, se prospectaron finalmente las 35 seleccionadas previamente por disponer de hábitats potenciales (figura 2).
Gracias al elevado número de participantes en el trabajo de campo se ha logrado una cobertura óptima en prácticamente toda el área de estudio. En casi todas las cuadrículas se pudieron llevar a cabo en marzo y abril el mínimo de muestreos requerido en la metodología propuesta (en varios casos se realizó un número considerablemente mayor). La única excepción ha sido la WN55, cuyos recorridos no pudieron efectuarse hasta primeros de mayo. En varias zonas, por otra parte, se ha extendido el periodo del trabajo de campo al mes de mayo, con el fin de confirmar la presencia del picamaderos en localidades con indicios y/o citas dudosas. Aunque en teoría fuera del periodo más adecuado para la detección de la especie, varios territorios se han confirmado gracias a las visitas efectuadas en estas fechas tardías.
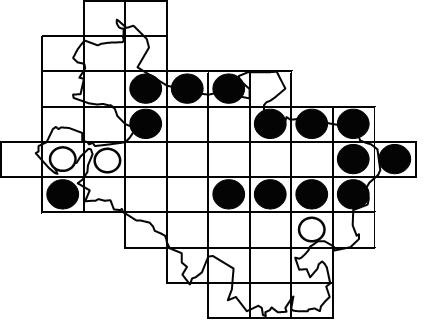
Los datos aportados por los participantes han permitido constatar fehacientemente -mediante la observación o escucha de ejemplares- la presencia del picamaderos negro en 14 de las cuadrículas estudiadas. En todas se han encontrado, además, rastros de alimentación inequívocamente atribuibles a este pícido. En tres cuadrículas más no ha sido posible registrar contactos directos, aunque se han hallado en ellas indicios sospechosos o se dispone de citas en época de reproducción anteriores a 2011 (figura 3).
Así pues, el área de distribución del picamaderos negro en Álava abarcaría en la actualidad el macizo de Gorbea y sus estribaciones; los montes de Arlabán; la vertiente meridional de las sierras de Elguea-Urkilla-Altzania desde Larrea hasta el límite con Navarra; la alineación formada por Entzia-Iturrieta-Montes de Vitoria hasta, aproximadamente, el monte Pagogan; y la sierra de Árcena, aunque en este caso la especie ha aparecido sólo marginalmente en la parte alavesa. Zonas donde existen sospechas de presencia, pero no ha podido ser confirmada por el momento, son los pinares de Valdegobía, el sector más occidental de Montes de Vitoria y la periferia de la depresión de Izki. Aunque existen algunas citas recientes, no se han obtenido indicios de ningún tipo en la comarca Cantábrica Alavesa-Orduña, ni en las sierras de Kodes y Lokiz. Por último, tampoco se han localizado indicios en los hayedos y pinares de la vertiente norte de la Sierra de Cantabria, y en este caso ni siquiera constan citas o avistamientos.
En cuanto a una aproximación al tamaño de población, se ha entendido que la detección de varios ejemplares en una misma jornada, o en jornadas separadas, podía interpretarse como la presencia de individuos territorializados. Analizando las distancias lineales entre diferentes contactos, y en función de los dominios vitales descritos para la especie en la bibliografía, puede estimarse provisionalmente entre 19 y 22 el número de “territorios” en el área de estudio. Los ejemplares asentados o territoriales no necesariamente son nidificantes, pero cuantificar esa fracción de la población ha estado fuera del alcance de este trabajo. Por otro lado, la falta de conocimientos sobre la autoecología de la especie en la Península Ibérica en general y en el área de estudio en particular impide saber si existe una población flotante o dispersante, que pudiera generar una sobrevaloración del número de territorios.
En conclusión, con este estudio se ha podido documentar la rápida colonización y asentamiento de una especie ausente del área de estudio hasta finales de los años 90. Las vías de expansión claramente se han focalizado a través de las sierras con mayor extensión de hayedos, aunque algunas formaciones de coníferas de montaña (pino albar y laricio), en buena parte plantaciones, también han sido ocupadas, al menos como zonas de alimentación. Las variaciones en la estructura y calidad de los rodales forestales, así como su proximidad geográfica y conectividad con las áreas previamente ocupadas por el picamaderos negro (áreas fuente) podrían explicar las diferencias en la abundancia de la especie, o su ausencia en determinadas comarcas.
Participantes voluntarios
G. Abascal, I. Agirre, D. Alday, R. Apaolaza, R. Arambarri, C. Arkotxa, G. Belamendia, E. Bernedo, J. Buesa, M. Corral, J. Echegaray, B. Fernández, J. M. Fernández (coordinador), J. A. Gainzarain (coordinador), J. Garayo, J. I. García, A. Gracianteparaluceta, M. Iglesias, S. de Juan, L. Lobo, J. Markinez, I. Martínez, J. L. Pina, P. Ruiz de Arkaute, N. Ruiz de Azua, M. Salvador, F. J. Sesma, F. Silván, A. Unanue y B. Valcárcel.
L. Abin, I. Ayala, E. Cabanillas, A. Cantero, M. Carrasco, L. Dueñas, J. Enjuto, J. M. Gómez, A. Ibáñez de Maeztu, L. Lacha, A. Montoya, R. Preciado, A. Puente, J. Rubines, M. Sáenz de Buruaga, A. Tirados, J. Villasante, A. Zufiaur y Consultora de Recursos Naturales SL.